- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2015
- Structure of materials
- The coelacanth out of breath
The coelacanth out of breath
Although it has been studied extensively, the extant coelacanth Latimeria still has some anatomical features that are not well understood. Based on propagation phase contrast X-ray microtomography performed at beamline ID19 as well as conventional tomography techniques, the development and evolutionary history of the coelacanth lung has been recently studied. The presence of a lung in Latimeria, its allometric growth through the ontogeny and the development of a fatty organ as an adaptation to deep-waters has been confirmed.
Coelacanths are a group of sarcopterygian fishes known since the Devonian, 410 million years (Ma) ago. Due to their absence from the record during the Cenozoic Era (65 Ma ago to present), they were thought to be extinct until the first specimen of the extant species Latimeria chalumnae [1] was caught in deep-waters of the Indian Ocean in 1938. This discovery has been the subject of many articles [1-4], thereby making the coelacanth one of the most famous large marine vertebrates. Among these studies, some [2, 4] have mentioned the presence of a vestigial lung apparatus in L. chalumnae, which merited further investigation.
The adult, juvenile and embryonic Latimeria specimens studied here belong to different institutional collections: South African Institute for Aquatic Biodiversity (Grahamstown, South Africa), Zoologische Staatssammlung (München, Germany) and Muséum National d’Histoire Naturelle (Paris, France). Direct observations on partial dissections and on histological material were made in the Collection of Comparative Anatomy of the MNHN.
The X-ray imaging study was mostly conducted at beamline ID19. Three embryos and a juvenile individual were scanned using long propagation (up to 13 metres) phase-contrast synchrotron X-ray microtomography. Two adult specimens were scanned in a hospital in Paris and on the AST-RX Platform of the MNHN, Paris, respectively.
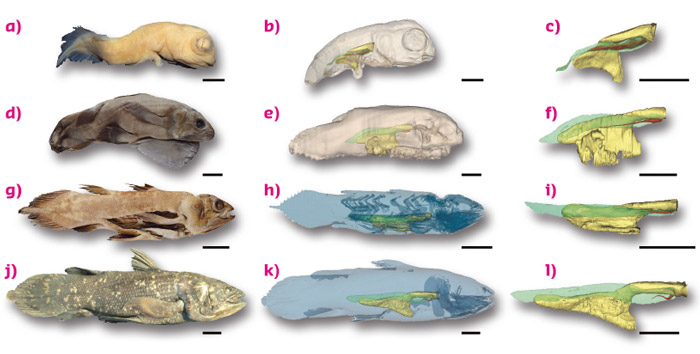
This study allowed us to confirm the presence of a non functional lung in extant coelacanths, and demonstrate an allometric growth of the lung relative to the adjacent correlated organs (digestive tract, fatty organ) during ontogenetic development (Figure 36). Although the growth of the fatty organ is correlated to the total length of the animal through the different ontogenetic stages, the lung presents a decreasing growth, exhibiting a vestigial shape at late ontogenetic stages. Due to the small number of invaluable coelacanth specimens housed in collections worldwide, and especially of embryos, access to high quality tomography facilities was necessary to study the fine anatomy of individuals of a unique ontogenetic series, without damage on these rare specimens. In this respect, the use of long propagation phase contrast made it possible to obtain high levels of contrast for soft tissues.
 |
|
Fig. 36: Photos and three-dimensional reconstructions of the lung and the fatty organ, in relation to the digestive tract, in a unique ontogenetic series of Latimeria chalumnae in right lateral views. (a, b, c) Early embryo CCC 202.1. (d, e, f) Late embryo with yolk sac CCC 29.5. (g, h, i) Juvenile CCC 94. (j, k, l) Adult CCC 22. Yellow, oesophagus and stomach; green, fatty organ; red, lung. Scale bar, 0.5 cm (a, b, c); 2.5 cm (d, e, f); 5 cm (g, h, i); 10 cm (j, k, l). |
Considering the structures of the coelacanth pulmonary apparatus through time, we postulate that adaptation to deep water led to the reduction of the lung, the complete loss of its respiratory function and the development of the fatty organ for buoyancy control. We also proposed that this adaptation - probably initiated by the end of the Mesozoic (100-65 Ma ago) - may have allowed the survival of deep-water living coelacanths during the Cretaceous/Paleogene extinction event.
In Latimeria, the fatty organ has a buoyancy function [5], possibly analogous to that of the liver of some sharks, which store lipids as a reserve, the fat-invested string of tissue of some deep-water actinopterygians fishes, and the spermaceti of some cetaceans, whose function is to maintain neutral buoyancy both at the surface and at depth.
Principal publication and authors
Allometric growth in the extant coelacanth lung during ontogenetic development, C. Cupello (a), P.M. Brito (a), M. Herbin (b), F.J. Meunier (c), P. Janvier (d), H. Dutel (e) and G. Clément (d), Nature Communications 6, 8222 (2015); doi: 10.1038/ncomms9222.
(a) Departamento de Zoologia, Universidade do Estado do Rio de Janeiro (Brazil)
(b) Département Écologie et Gestion de la Biodiversité, UMR 7179 CNRS–MNHN, Muséum National d’Histoire Naturelle, Paris (France)
(c) Département des Milieux et Peuplements Aquatiques, UMR 7208 (CNRS–IRD–MNHN–UPMC), Muséum National d’Histoire Naturelle, Paris (France)
(d) Centre de Recherche sur la Paléobiodiversité et les Paléoenvironnements (CR2P, UMR 7207), Sorbonne Universités, MNHN, CNRS, UPMC-Paris6, Muséum National d’Histoire Naturelle, Paris (France)
(e) Laboratory for Evolutionary Morphology, RIKEN, Kobe (Japan)
References
[1] J.L.B. Smith, Nature 143, 455–456 (1939).
[2] J. Millot, J. Anthony, D. Robineau, in Anatomie de Latimeria chalumnae III (CNRS, 1978).
[3] C.T. Amemiya et al., Nature 496, 311–316 (2013).
[4] D. Robineau, Ann. Sci. Nat. Zool. 8, 43–60 (1986).
[5] P.M. Brito et al., Palaeontology 53, 1281–1290 (2010).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.