- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2009
- X-ray imaging
- Untangling the evolution of giant sperm in ostracod microcrustaceans using holotomography
Untangling the evolution of giant sperm in ostracod microcrustaceans using holotomography
Some animal groups have extraordinarily long and thin filamentous spermatozoa. This characteristic usually occurs only in a single or just a few species up to a subgenus like in Drosophila. However, there is one exception: ostracods, which are micro-crustaceans living worldwide in all kinds of water bodies. Within their superfamily Cypridoidea, all circa 1600 living, mainly freshwater, species show reproduction with giant sperm (sperm length up to 7 times the body length in Heterocypris sydneia). The fact that giant sperm are present in the whole taxon suggests that this way of reproduction developed in the early phase of the evolution of this group.
In addition to giant sperm, an intricate set of reproductive organs developed in this group. Both genders have a complete system of reproductive organs on either side of the body. Males possess a paired sperm pump (or Zenker organ) within the sperm ducts, which is mainly a sclerotised chitinous tubular structure. Two vaginas of the female accommodate the two penes of the male during copulation to receive the giant sperm, which are then transferred to a pair of storage organs (or seminal receptacles), before they are used for egg fertilisation.
Ostracods are the most abundantly preserved arthropods in the fossil record dating from at least the Ordovician (450 Myr). Their exceptional fossil record is due to their calcitic bivalved carapace, usually 0.3 to 3 mm long, that is readily preserved. Unlike the carapace, the body and appendages are only very rarely recorded as fossils, and mostly with only a poor preservation of the appendages. An exception to this is Harbinia micropapillosa from the Santana Formation of the Cretaceous of Brazil (about 100 million years old). Specimens of this species are hosted in the Natural History Museum’s collection in London (NHM). Previous scanning electron microscopic studies of this species showed that the external surface of the soft parts is frequently preserved in very high quality by apatite. This present study reaveals the internal features for the first time.
Of 23 specimens of H. micropapillosa (a loan of the NHM), and 18 specimens of the living Eucypris virens (used as comparative material), holotomographic scans were performed at beamline ID19 with a monochromatic beam at an energy of 20 keV and a detector pixel size of 0.678 micrometres. Each holotomographic data set was composed of four different scans using 1500 projections over 180 degrees: one absorption scan (sample to detector distance of 5 mm) and three propagation phase contrast scans at increasing distances (20 mm, 40 mm, 70 mm). Phase retrieval was computed for each projection angle with an algorithmic approach adapted to absorbing samples. Reconstructed holotomographic slices of fossil specimens were then optimised by attenuating the most contrasted parts (mostly due to pyrite infilling) and by normalising the air background.
Each fossil specimen of less than 1 mm body length was finally represented by a stack of 1200 to 1500 virtual slices, which were further processed within 3D-visualising software.
 |
|
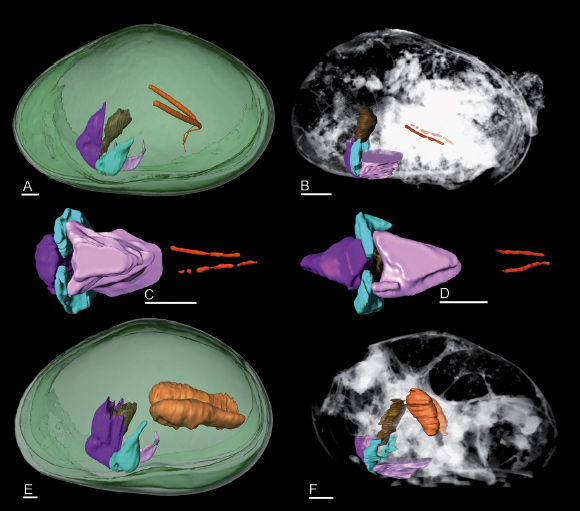
Fig. 144: Reproduction organs in Eucypris virens (extant) and Harbinia micropapillosa (fossil). Anterior to the left. (A and E) lateral view of male (A) and female (B) E. virens with several organs shown for comparison. (B and F) lateral view of male (B) and female (F) H. micropapillosa with several organs shown in context of whole body reconstruction. (C and D) ventral views of several organs including tubes of Zenker organs of male H. micropapillosa. Colours: orange: central tubes of Zenker organs in males, seminal receptacles in females; brown: oesophagus; turquoise: mandible; purple: upper lip; pink: lower lip; green: valves; grey-scales: whole body reconstruction. All scale bars 100 µm. Original data underlying the cited publication can be downloaded from the ESRF Palaeontological Microtomographic Database (http://paleo.esrf.eu). |
Processing of the holotomographic data obtained on the fossil specimens of H. micropapillosa revealed structures of biological origin within the main body of the animals (Figure 144). In males, a pair of tubes close to the penes can be identified as traces of Zenker organs, while large paired cavities near the mid-point of the body represent the seminal receptacles in female specimens. The receptacles must have been filled with sperm in order to be preserved as two cavities – rare fossil evidence of insemination. Since both Zenker organs and large seminal receptacles are only known from ostracods reproducing with giant sperm, this extraordinary method of reproduction must have developed in this group of ostracods at least 100 million years ago. The conformity of fossil and recent reproductive organs suggest that giant sperm probably developed only once in this group.
Thanks to the outstanding resolution and contrast features of holotomography at the ESRF we found that in freshwater ostracods, reproduction with giant sperm has been a successful and stable feature for a long time – despite high biological costs. Exploring the advantages of this exceptional kind of reproduction will be the focus of our ongoing research.
Principal publication and authors
R. Matzke-Karasz (a), R.J. Smith (b), R. Symonova (c), C.G. Miller (d) and P. Tafforeau (e), Science 324, 1535 (2009).
(a) Ludwig-Maximilians-University Munich (Germany)
(b) Lake Biwa Museum, Kusatsu (Japan)
(c) Charles University, Prague (Czech Republic)
(d) Natural History Museum London (UK)
(e) ESRF
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.