- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2013
- X-ray imaging
- Hearing by word of mouth
Hearing by word of mouth
X-rays enabled the discovery of a new hearing mechanism for animals lacking an ear. Using X-ray synchrotron holotomography, the biomechanical properties of ear tissues have been measured and a model of the acoustic propagation and resonator mode generated from these measurements.
The mechanism of sound perception is globally well understood. We know that the outer ear receives sound waves and that the tympanic or middle ear transmits these waves to the eardrum. The sound waves make the eardrum vibrate, and the eardrum delivers these vibrations to the inner ear using one or several ossicles. Finally specialised nerve cells of the inner ear translate the vibrations into nervous pulses sent to the brain.
Vertebrates such as mammals, birds, crocodylids, turtles and squamatas use this mechanism. However, we know that the middle ear evolved independently many times in terrestrial tetrapods [1]. The functional anatomy of hearing in frogs differs from that of other terrestrial tetrapods. Most frogs have only a tympanum at the head surface, even though two species use ultrasound call and are equipped with a tympanic cavity [2,3,4]. Moreover, several species are “earless” in the sense that they lack a tympanic middle ear. Indeed, we know that approximately three hundred frog species lack a tympanic middle ear.
Gardiner’s frogs (Sechellophryne gardineri) from the Seychelles are one of the smallest frogs in the world with a length of about 10 mm (Figure 17). They do not possess a middle ear with an eardrum yet they produce acoustic signals. These frogs are members of the Sooglossidae family and they evolved in isolation on the Seychelles Archipelago over the last 47–65 My. We sought to understand how these frogs perceive sound without a middle ear and eardrum, and how they perceive sound when 99.9% of any sound wave reaching an animal is reflected at the surface of its skin.
 |
|
Fig. 17: Gardiner’s frog in natura and volume rendering obtained by a holotomography scan. |
To investigate whether Gardiner frog’s use sound to communicate with each other, we set up loudspeakers in their natural habitat to broadcast an array of pre-recorded mating calls. This caused male frogs present in the rainforest to answer. Therefore we established that Gardiner frogs are able to hear and that they use acoustic communication. The next step was to understand the hearing mechanism used by this species of apparently deaf frogs, from an anatomical point of view.
Various mechanisms have been proposed for other frogs without a middle ear: an extra-tympanic pathway through the lungs, muscles connected to the inner ear, and bone conduction. We used X-ray tomographic techniques at beamline ID19, including holotomography, to scan Gardiner’s frogs and related frogs equipped with a middle ear (Figure 18). We then constructed 3D models of these frogs to analyse the head’s functional anatomy in detail. We established that neither the pulmonary system nor the muscles could contribute significantly to the transmission of sound to the inner ears.
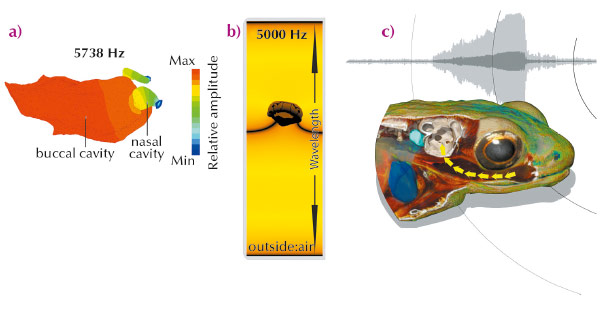 |
|
Fig. 18: a) Numerical simulation of bone conduction and mouth cavity (resonator). b) Volume rendering of head illustrating the sound transmission in to inner ear. c) Illustration of the Gardiner’s frog hearing mechanism: the mouth acts as a resonating cavity for the frequencies of the frog’s song, amplifying the amplitude of the sound in the mouth. The body tissue between the buccal cavity and the inner ear is specially adapted to transmit these sound waves to the inner ear. |
We then used numerical simulations to investigate the possibility that the sound was received through the frog’s head (Figure 18). These simulations highlighted the key role played by the mouth which acts as a resonator, or amplifier, for the frequencies emitted by this species (Figure 18). X-ray imaging of different species showed that the transmission of the sound from the oral cavity to the inner ear has been optimised by two evolutionary adaptations located between the mouth and the inner ear, a reduction in thickness of the tissue and a smaller number of tissue layers. Our conclusion is that sound perception in Gardiner’s frogs is without use of a tympanic middle ear and occurs by the unique combination of a resonating mouth cavity and bone conduction (Figure 18).
Principal publication and authors
R. Boistel (a,b,c), T. Aubin (a,b), P. Cloetens (d), F. Peyrin (d,e), T. Scotti (f), P. Herzog (f), J. Gerlach (g), N. Pollet (h), and J.-F. Aubry (i), Proc Natl Acad Sci U S A. 110, 15360-15364 (2013).
(a) Centre de Neurosciences Paris-Sud (CNPS), Centre National de la Recherche Scientifique (CNRS) Unité Mixte de Recherche (UMR) 8195, Université Paris XI, Orsay (France)
(b) Université Paris Sud, Orsay (France)
(c) Institut International de Paléoprimatologie et de Paléontologie Humaine (IPHEP), CNRS UMR 7262, Université de Poitiers (France)
(d) ESRF
(e) Centre de Recherche en Acquisition et Traitement de l’Image pour la Santé, CNRS UMR 5515, Institut national des sciences appliquées de Lyon, Villeurbanne (France)
(f) Laboratoire de Mécanique et d’Acoustique (LMA), CNRS UPR 7051, Marseille (France)
(g) Nature Protection Trust of Seychelles, Victoria, Mahé (Seychelles)
(h) Institute of Systems and Synthetic Biology, Genopole, CNRS, Université d’Evry Val d’Essonne (France)
(i) Institut Langevin, CNRS UMR 7587, École supérieure de physique et de chimie industrielles de la ville de Paris ParisTech (France)
References
[1] J.W.H. Schnupp and C.E. Carr, Nat Neurosci 12, 692–697 (2009).
[2] A.S. Feng, P.M. Narins, C.H. Xu, W.Y. Lin, Z.L. Yu, Q. Qiu, Z.M. Xu and J.X. Shen, Nature 440, 333-6 (2006).
[3] V.S. Arch, T.U. Grafe, M. Gridi-Papp and P.M. Narins, PLoS One 4, e5413 (2009);
[4] R. Boistel et al. PLoS One 6, e22080 (2011).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.