- Home
- News
- Spotlight on Science
- Beating heart efficiency...
Beating heart efficiency revealed by X-rays
10-04-2017
The heart pumps blood through the vessels while keeping the pulmonary and systemic circulatory systems in balance. At the molecular level, this balance is controlled by mechanosensing in the myosin filaments that recruit myosin motors in relation to the load.
The contraction of the heart, like that of skeletal muscle, is due to the reciprocal sliding of two kinds of overlapping protein filaments, the thin actin-containing filament and the thick myosin-containing filament [1, see also: Small angle X-ray diffraction reveals muscle’s gearbox]. These filaments are located within each sarcomere, which is the 2 µm long structural unit repeating over the whole length of the muscle cell. A globular portion of the myosin molecule, protruding from the thick filament, is the molecular motor that generates force and muscle shortening by pulling the thin filament towards the centre of the sarcomere during cyclic ATP-driven interactions. In the heart, an intrinsic regulatory mechanism allows the force developed by the ventricle wall during the contraction phase (systole) to be adjusted with respect to its filling during the preceding relaxation phase (diastole), ensuring the balance between the two circulatory systems, pulmonary and systemic, working in series. This process is described by the Frank-Starling law, which dates from 1918. At the cell level, the Frank-Starling law implies that the contractile force increases with the increase in the sarcomere length.
In this work, we demonstrate that the molecular basis of the Frank-Starling law relies on a mechanosensing-based regulation of thick filament activation: in the resting state, the myosin motors lie in their off state on the surface of the thick filament, unable to attach to actin and split ATP; during contraction, the number of motors leaving the resting state is adjusted to the force required to pump blood into the aorta or the pulmonary artery by mechanosensing in the thick filament. In this way, the energetic cost of the contraction is tuned to the mechanical task.
Intact trabeculae, pillars of muscle cells 2-3 mm in length (Figure 1a), were mounted at beamline ID02 between force and length transducers to record together the mechanical responses and the X-ray diffraction signals from the myosin motors during electrically-paced contraction-relaxation cycles at different sarcomere lengths (Figure 1b,c). Sub-nanometre precision in the measurement of axial movements of the myosin motors was attained by exploiting the interference fine structure in the low-angle X-diffraction pattern from the bipolar arrays of myosin motors in each sarcomere. The combination of sarcomere-level mechanics [2] and USAXS/SAXS at ID02 in intact trabeculae made it feasible to record the nanometre-scale signals from the contractile proteins along the myosin and actin filaments together with the micrometre-scale changes in the length of the sarcomere (Figure 2a-d).
 |
|
Figure 1. a) Photograph of the internal wall of the heart showing 2-3 mm long trabeculae projecting as pillars from the inner surface of the right ventricle. The photograph was taken just before dissection of the central trabecula (arrow). b) Force of the FE (red) and LC (green) twitches. Stimulus starts at 775 ms. Grey bars: X-ray time windows. c) Relation between the peak force and the sarcomere length (dashed line) superimposed on the instantaneous force-sarcomere length plots for twitches in FE (red) and in LC (green). Filled circles, peak forceaccording to the same colour code as the traces. The shaded areas are the two ideal loops followed by contractions that start at sarcomere length ~2.2 µm under isometric condition and become isotonic when the force attains the same level as the twitch peak. In terms of the of the left ventricle beat, the dashed line is the relation between end-systolic pressure and end-systolic volume and the dashed areas are two pressure-volume loops starting from the same preload (i.e. end-diastolic volume) and with two different aortic pressures. |
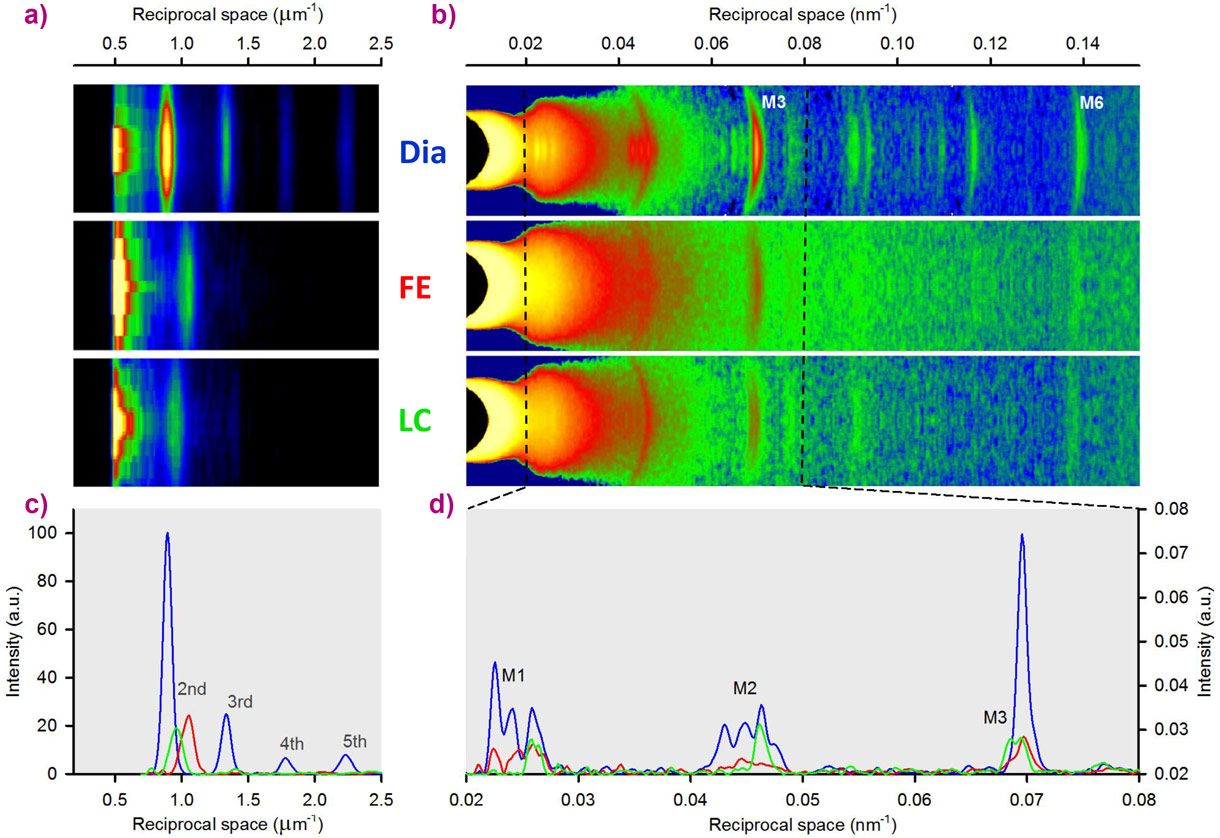
During force development under fixed-end conditions (FE), sarcomeres shorten against the end compliance so that, at peak force, sarcomere length reduces from the starting value of 2.21 ± 0.02 µm (Figure 2a,c, blue) to 1.9 ± 0.03 µm (red). In the subsequent twitch, further shortening is largely prevented (length clamp conditions, LC), by a feedforward signal proportional to the shortening in the preceding FE twitch, so that, at peak force, sarcomere length is 2.09 ± 0.01 µm (green) and the force is twice the FE value (Figure 1b,c). We find that in diastole (Dia, Figure 2b,d, blue) all the myosin-based reflections mark the quasi-helical three-stranded symmetry followed by the myosin molecules when they lie in their off state on the surface of the thick filament, unable to attach to actin and split ATP [1,3,4]. At the peak of the twitch force (red and green) the intensities of all the meridional reflections decrease, due to the myosin motors leaving their helical tracks as the thick filament switches on. The M3 reflection, originating from the axial repeat of the myosin motors, exhibits a different fine structure and different increase in spacing depending on whether the contraction occurs in FE (red) or in LC (green). In terms of the structural model of the sarcomere [5], the difference in M3 intensity profiles indicate that, during a cardiac twitch, only a fraction of the motors leave the off state and this fraction depends on the level of the force independently of the diastolic sarcomere length. In this respect, the LC and FE twitches approximate the conditions of the left ventricle beating against a high (LC) and a low (FE) aortic pressure. In turn, the end-systolic pressure-volume relation of the left ventricle is the organ correlate of the active tension-length relation determined at sarcomere level with twitches that, according to our protocol, start from the same length and experience different loading conditions (Figure 1c).
 |
|
Figure 2. a) Meridional slices of X-ray patterns collected at 30 m from the preparation during diastole (Dia) and at the peak force of a FE or LC twitch, showing the first orders of sarcomeric reflections. Total exposure time 20 ms for Dia, 10 ms for FE and LC. b) Meridional slices of patterns collected at 1.6 m from the preparation, showing the myosin-based axial reflections (M1-M6). Total exposure time 150 ms for Dia, 60 ms for FE, 90 ms for LC. c) Superimposed intensity profiles from (a), starting from the second-order reflection. d) Superimposed intensity profiles from the region of (b) indicated by the dashed lines. |
This work provides a new integrated view of the Frank-Starling mechanism, which combines mechanical, structural, and energetic aspects: independent of the end-diastolic sarcomere length (end-diastolic volume), the energetic cost of the heart beat is tuned to the ventricular end-systolic pressure-volume relation by stress-sensing in the thick filament. Moreover, the work establishes a new methodology based on the combination of sarcomere-level mechanics with nanometre-micrometre scale X-ray diffraction on heart cell bundles from biopsies and explants for both the investigation of how cardiomyopathy-causing mutations impair heart function and the testing of specific small molecule effectors for new therapeutic interventions.
Principal publication and authors
Myosin filament activation in the heart is tuned to the mechanical task, M. Reconditi (a,b), M. Caremani (a), F. Pinzauti (a), J.D. Powers (a), T. Narayanan (c), G.J.M. Stienen (d), M. Linari (a,b), V. Lombardi (a), and G. Piazzesi (a), PNAS 114, 3240-3245 (2017); doi: 10.1073/pnas.1619484114.
(a) PhysioLab, University of Florence (Italy)
(b) Consorzio Nazionale Interuniversitario per le Scienze Fisiche della Materia, Unità di Ricerca di Firenze, Florence (Italy)
(c) ESRF
(d) Department of Physiology, VU University Medical Center, Amsterdam (The Netherlands)
References
[1] Linari et al., Nature 528, 276 (2015).
[2] Caremani et al., PNAS 113, 3675 (2016).
[3] Zoghbi et al., PNAS 105, 2386 (2008).
[4] Stewart et al., PNAS 107, 430 (2010).
[5] Reconditi et al., PNAS 108, 7236 (2011).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.