- Home
- News
- Spotlight on Science
- How prions are generated...
How prions are generated by a copper switch
09-12-2015
The conversion of the prion protein into a misfolded isoform in the brain causes prion diseases, a group of transmissible neurodegenerative disorders affecting humans and animals. The structural events underlying this conversion have escaped detailed characterisation due to the insoluble nature of prions. By using XAS at beamline BM30B, a team of Italian scientists have identified a novel copper-mediated mechanism that acts as a switch that turns the normal protein into its pathological alter ego.
Prions are misfolded proteins that cause a group of incurable neurodegenerative diseases, including spongiform encephalopathies, such as mad cow diseases and Creutzfeldt-Jakob disease in human. Prions are unique infective agents —unlike viruses, bacteria, fungi and other parasites, prions do not contain either DNA or RNA. They can propagate their pathological effects like “wildfire” by infecting the normal proteins (termed PrPC) causing them to acquire the wrong conformation and convert into further disease-causing agents called prions or PrPSc. Despite the importance for pathogenesis, the mechanism of prion formation has escaped detailed characterisation due to the insoluble nature of prions. PrPC interacts with copper through octarepeat and non-octarepeat binding sites. Copper coordination to the non-octarepeat (non-OR) region has garnered interest due to the possibility that this interaction may impact prion conversion.
We have identified a new mechanism underlying the pathological transformation of prion proteins: it all depends on a metal, copper, and its bond with the protein. We used extended X-ray absorption fine structure (EXAFS) spectroscopy, cell-biology and molecular dynamic simulations to investigate how copper coordination in the fifth copper binding site may influence prion conversion.
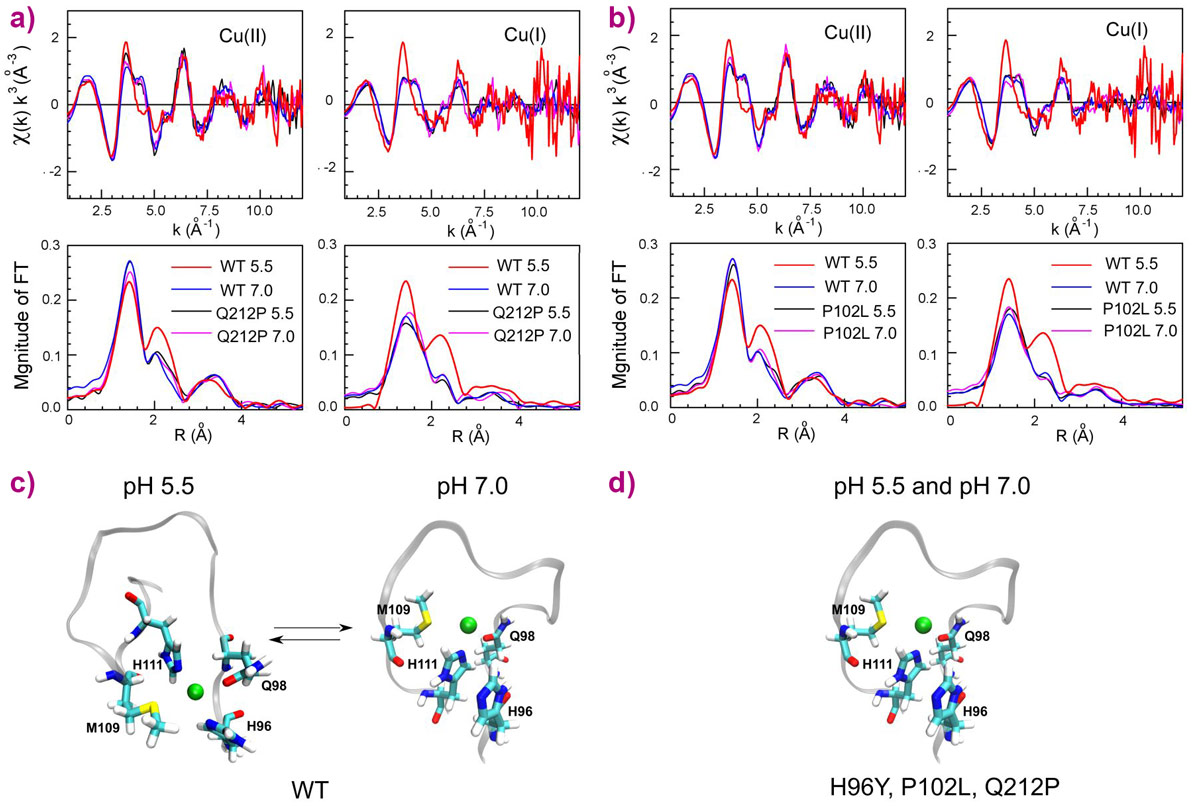
Using X-ray absorption spectra collected at beamline BM30B FAME, we determined the copper coordination in the wild type (WT) human prion protein (HuPrP) and in different pathological mutants at both pH 7.0 and pH 5.5. We found that mutations (H96Y, P102L and Q212P) and pH changes cause a dramatic modification on both Cu(II) and Cu(I) coordination in the non-OR region. In the WT HuPrP Cu(II) and Cu(I) are anchored to His96 and His111 only at pH 5.5, while at pH 7.0 copper at both oxidative states is coordinated by His111. Conversely, in the mutants copper is bound only to H111 independently of the pH (Figure 1).
 |
|
Figure 1. Copper coordination in the WT and mutant human prion protein. k3-weighted EXAFS spectra and Fourier transforms of Cu(II) and Cu(I) bound to WT HuPrP(90–231) and Q212P at pH 5.5 and 7.0 (a) and of Cu(II) and Cu(I) bound to WT HuPrP(90–231) and P102L at pH 5.5 and 7.0 (b). Schematic representations of copper binding sites in the WT HuPrP(90–231) (c) and in the mutants (d) at both pH 5.5 and 7.0. |
The observed structural differences in the copper coordination among WT and pathological mutants at pH 5.5 may have relevant physiological effects since this alteration in the copper binding site might trigger PrPC to PrPSc conversion. To understand the physiological implications of EXAFS data, we performed in vitro and cell-based assays. Intriguingly, the removal of one single copper ligand (H96) promoted prion conversion and generation of infectious prion material in neuronal cells.
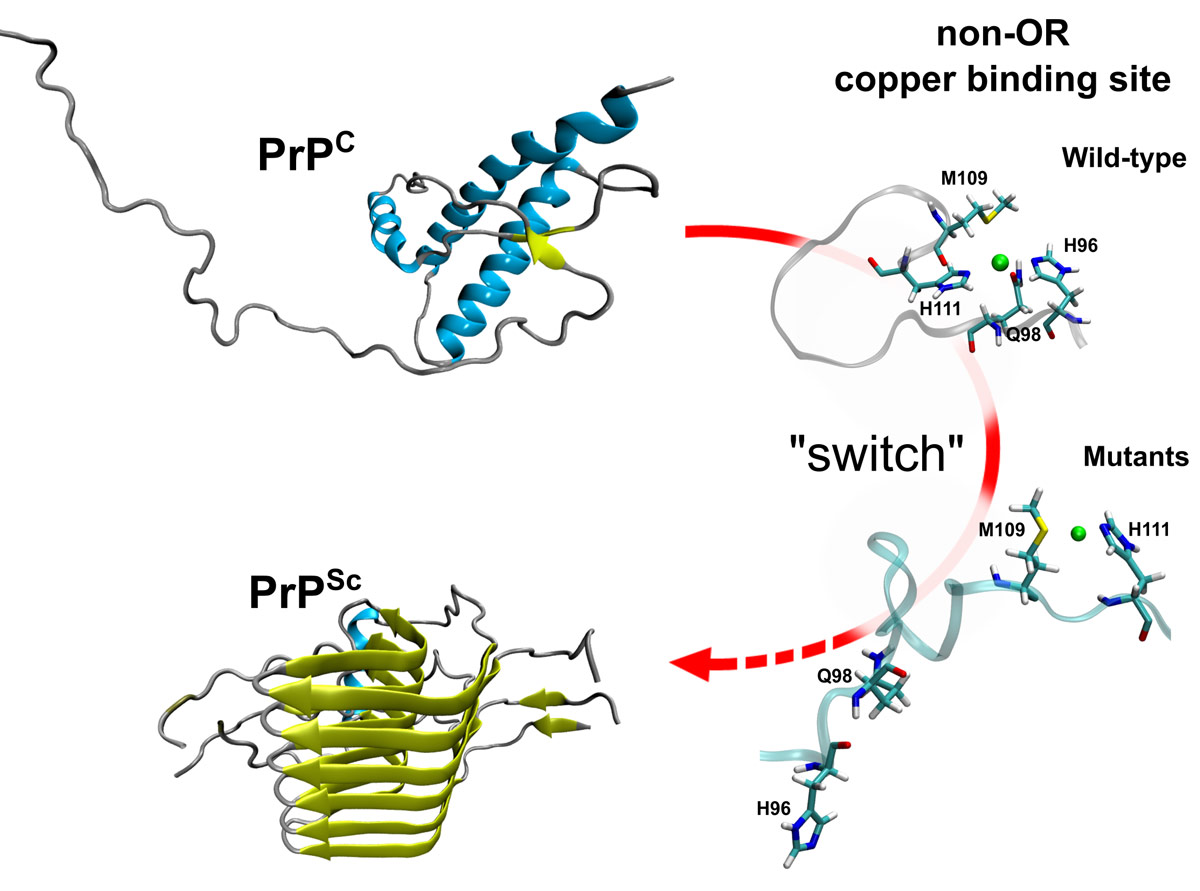
These findings suggest a pivotal role for non-OR region as a critical molecular switch for prion conversion. We therefore argue that copper bound to the non-OR region may stabilise this segment when coordinated by His96 and His111, preventing misfolding events through transient short and long range interaction contacts between the 90–127 residues and the C-terminal structured domain. This study highlights the importance of the non-OR region for prion conversion and suggests a model in which PrPC coordinating copper with one His may be more prone to the conversion in acidic conditions (Figure 2).
 |
|
Figure 2. Model for the non-OR region molecular switch at acidic pH. PrPC coordinating copper with one His residue in the non-OR region is more prone to the conversion at acidic pH condition. We propose a model where His96 and His111 represent the N-terminal switch for prion conversion in the PrPC. |
Principal publication and authors
G. Giachin (a,b), P.T. Mai (b), T.H. Tran (b), G. Salzano (b), F. Benetti (b), V. Migliorati (c), A. Arcovito (d), S.D. Longa (e), G. Mancini (f, g), P. D'Angelo (c), G. Legname (b, h), The non-octarepeat copper binding site of the prion protein is a key regulator of prion conversion, Sci Rep. 5, 15253 (2015); doi: 10.1038/srep15253.
(a) ESRF
(b) Department of Neuroscience, Scuola Internazionale Superiore di Studi Avanzati (SISSA), Trieste (Italy)
(c) Department of Chemistry, Sapienza University of Rome (Italy)
(d) Istituto di Biochimica e Biochimica Clinica, Università Cattolica del Sacro Cuore, Rome (Italy)
(e) Department of Medicine, Public Health, Life and Environmental Science, University of L’Aquila, Coppito Aquila (Italy)
(f) Scuola Normale Superiore, Pisa (Italy)
(g) Istituto Nazionale di Fisica Nucleare (INFN) sezione di Pisa (Italy)
(h) ELETTRA - Sincrotrone Trieste S.C.p.A (Italy)
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.