- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2017
- Structure of materials
- Sponge-associated bacteria mineralise arsenic and barium on intracellular vesicles
Sponge-associated bacteria mineralise arsenic and barium on intracellular vesicles
Arsenic and barium are ubiquitous and toxic elements. If accumulated by organisms from their environment, they are excreted, assimilated into organic forms or mineralised. This work demonstrates that the bacterium Entotheonella sp. found in the Red Sea sponge Theonella swinhoei is capable of intracellular biomineralisation of both elements, on membrane vesicles.
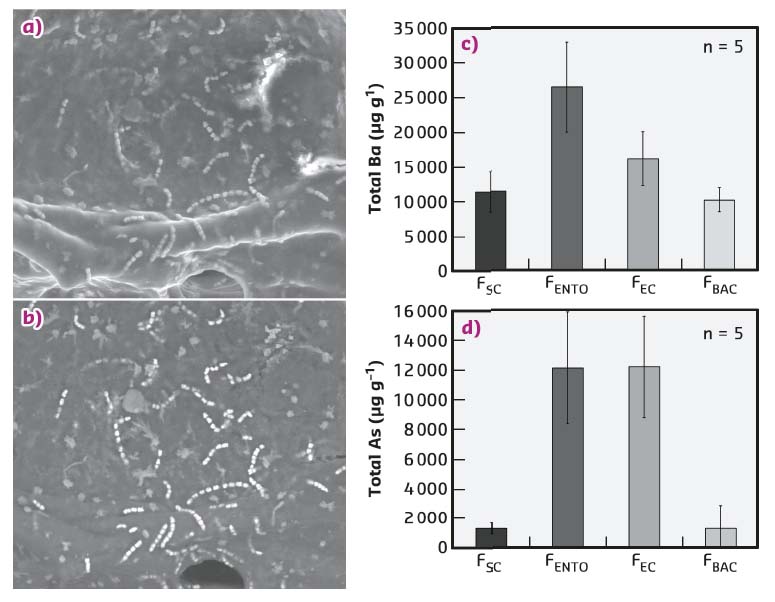
Previous research found hyper-accumulation of arsenic and barium in the Red Sea sponge, T. swinhoei [1]. Fractionation of T. swinhoei cells and the ICP-MS elemental analysis of the resulting fractions showed that the filamentous bacteria Entotheonella sp. constitutes the arsenic and barium accumulating entity within its host (Figure 124). Entotheonella sp. is a filamentous bacterium described as trichomes or as chains of 4 to 20 cells. It is the most abundant bacteria found in T. swinhoei, comprising 3.25% of the sponge’s volume.
 |
|
Fig. 124: SEM micrographs of Entotheonella sp. in sponge tissue. a) Secondary electron detection mode indicating some of the Entotheonella sp. filaments. Scale bar, 100 µm. b) Backscattered electron detection mode reveals Entotheonella sp. filaments as the most electron-dense objects in the sponge. Scale bar, 50 µm. c, d) Dry weight concentration (µg g–1) of barium and arsenic (respectively) in cell-enriched fractions (n = 5). FBAC, unicellular bacterial cells; FEC, Entotheonella sp. with cyanobacteria; FENTO, Entotheonella sp. cells; FSC, sponge cells. Error bars show s.e. |
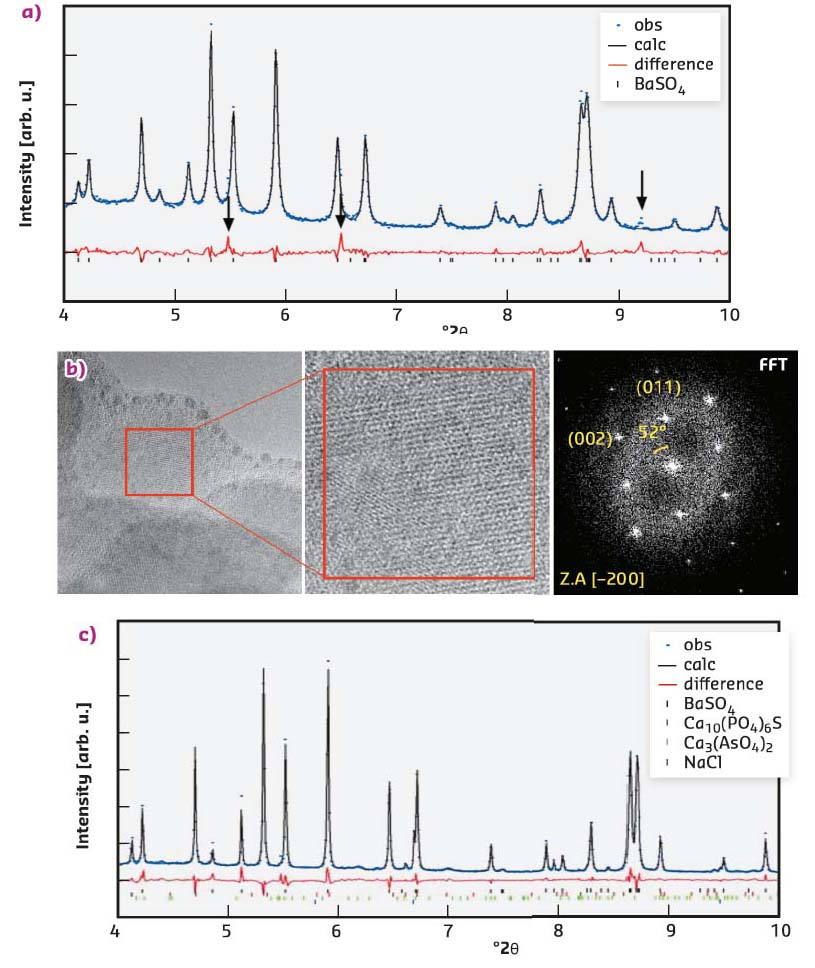
SEM scans and EDS analysis of the Entotheonella sp. filaments show they contain multiple barium- and arsenic-rich spherical granules inside their cytoplasm. The average sphere volume is 0.11 ± 0.007 µm3 (n = 121). The spheres have an organic core and are covered by an organic veneer. High-resolution synchrotron X-ray powder diffraction of freeze-dried samples obtained on ID22 showed that barite (BaSO4) was the major phase and with arsenates and phosphates as minor phases in the crystalline fraction (Figure 125a). Crystalline barite presence within spheres was also confirmed by high-resolution TEM examination of FIB-prepared thin section. The fast Fourier transform (FFT) of the TEM image obtained from a magnification close to an individual atomic lattice layer exhibits single crystal electron diffraction from the zone axis of barite (Figure 125b). Precise structural determination of the minor phases was achieved after mild thermal annealing. The resulting diffractogram clearly detects crystalline calcium arsenate (Ca3(AsO4)2) and calcium sulfide phosphate (Ca10(PO4)6S) as minor phases (Figure 125c).
 |
|
Fig. 125: XRD and TEM analysis of spheres inside Entotheonella sp. a) Rietveld refinement plot of preheated, freeze-dried Entotheonella sp. showing fit for observed (blue dots) and calculated (black line) diffraction patterns and the difference between them (red line). Black notches indicate the positions of the diffraction peaks of the major crystalline phase, barite (BaSO4). Diffraction peaks highlighted by arrows in the difference curve are compatible with an arsenate or phosphate phase. b) HRTEM micrograph of a portion of the freeze-dried Entotheonella sp. Insert shows higher magnification image revealing the lattice, and with its fast Fourier transform (FFT). c) Rietveld refinement plot of a freeze-dried Entotheonella sp. after thermal annealing: minor phases of sodium chloride (NaCl), calcium arsenate (Ca3(AsO4)2) and calcium sulfide phosphate (Ca10(PO4)6S). Observed (Obs.), calculated (Calc.) and difference curves are presented in the same schematics as in (a). |
The presence of barite and calcium arsenate were further confirmed on unprocessed flash-frozen Entotheonella sp. samples by synchrotron X-ray microprobe analyses performed at 95 K. The distribution of arsenic in the filaments was evident by micro-focused X-ray fluorescence (µXRF) mapping. All arsenic K edge X-ray absorption near edge structure (XANES) spectra of Entotheonella sp. showed As(V) as the dominant valence. Least-square linear combination fitting (LCF) of filament spectra to arsenic reference compounds was performed. Best fits were obtained using combinations of non-sulfur As(III), calcium arsenates (mostly pharmacolite) and sodium arsenate. µXRD performed on some of the filaments’ XANES locations showed the presence of pharmacolite.
These results show extraordinary concentrations of barium and arsenic in Entotheonella sp. (12,000 µg g-1 As is among the highest measured in any known organism). Entotheonella sp. is the only known prokaryote capable of intracellular barite mineralisation and only one other prokaryote, Desulfotomaculum auripigmentum, is known to intracellularly mineralise arsenic [2]. This is the first report of crystalline calcium arsenate, pharmacolite, as a biomineral. It is only in Entotheonella sp. that these elements are sequestered in a membrane-bound, organelle-like structure within the cytoplasm, and mineralised with many of the characteristics of controlled biomineralisation.
Principal publication and authors
Sponge-associated bacteria mineralize arsenic and barium on intracellular vesicles, R. Keren (a), B. Mayzel (a), A. Lavy (a), I. Polishchuk (b), D. Levy (b), S.C. Fakra (c), B. Pokroy (b) and M. Ilan (a), Nat Commun. 8, 14393 (2017); doi: 10.1038/ncomms14393.
(a) Department of Zoology, George S. Wise Faculty of Life Sciences, Tel Aviv University (Israel).
(b) Faculty of Materials Engineering and the Russell Berrie Nanotechnology Institute, Technion, Israel Institute of Technology (Israel).
(c) Advanced Light Source, Lawrence Berkeley National Lab, Berkeley (USA)
References
[1] B. Mayzel et al., PloS one 9, e95775 (2014).
[2] D.K. Newman et al., Applied and Environmental Microbiology 63, 2022-2028 (1997).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.