- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2017
- Structural biology
- Condensin complexes fasten the belt around DNA
Condensin complexes fasten the belt around DNA
The crystal structure of the major DNA binding site in the eukaryotic condensin complex, which is jointly formed by the Cnd3 HEAT-repeat and Cnd2 kleisin subunits, has been determined. The DNA-bound structure indicates an unconventional 'safety belt' mechanism that anchors condensin complexes and might provide condensin with the means to form large chromatin loops.
Ever since the first observation of condensed chromosomes in dividing cells during the late 19th century scientists have been trying to understand the principles that underlie this dramatic reorganisation of chromatin fibres [1]. However, it took more than one hundred years before the five-subunit condensin complex was identified as the leading player in the formation of mitotic chromosomes [2]. Thus, despite its key role in the correct segregation of chromosomes during mitosis and meiosis, the molecular mechanisms by which condensin complexes interact with chromatin fibres and alter their topology have, so far, remained largely mysterious.
Condensin complexes share their basic architecture with other structural maintenance of chromosomes (SMC) complexes, including cohesin and SMC5/6 in eukaryotes, and different SMC-type complexes in prokaryotes. All SMC complexes play essential roles in the maintenance of a cell’s genetic material during various aspects of chromosome biology, including chromosome condensation and segregation, DNA recombination and damage repair, and the organisation of chromosomal territories. The conserved basic architecture of these complexes suggests that they share a common mode of action in how they perform this array of seemingly diverse tasks.
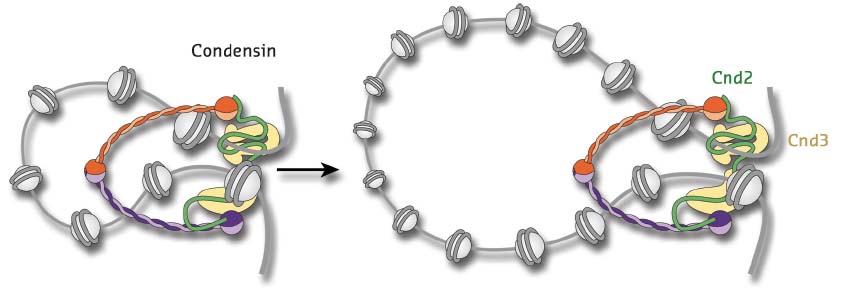
How would such a common mechanism work? A number of different models have been suggested over the years, but recently a process called ‘loop extrusion’ has been championed as a simple and elegant mechanism that could explain most of the cellular tasks of SMC complexes (Figure 11). How these complexes might form and extend DNA loops is, however, not understood, but one precondition of any loop extrusion model is that the proteins would need to make direct contact with their DNA substrates. Since all subunits of SMC complexes are largely devoid of classic DNA-binding motifs, the nature and specificity of this interaction has remained uncertain.
 |
|
Fig. 11: Model for the extrusion of chromatin loops by the five-subunit condensin complex. The complex is thought to encircle DNA within the large ring-shaped architecture formed by its Smc2-Smc4 coiled coil subunits (orange and purple) and Cnd2 kleisin subunit (green). HEAT-repeat subunit Cnd3 (yellow) binds to the Cnd2 kleisin. |
Previous work had demonstrated that a ternary sub-complex consisting of two HEAT-repeat subunits and the so-called kleisin subunit can facilitate DNA binding in a sequence-unspecific manner [3]. To identify the DNA binding site in this ternary sub-complex, the three subunits were expressed and purified individually and tested alone or in combination for DNA binding. Neither subunit alone could bind DNA efficiently. Only the combination of the HEAT-repeat protein Cnd3 with the kleisin Cnd2 yielded a DNA-binding activity comparable to the ternary complex.
This complex was – after some additional engineering and screening – amenable to crystallisation by vapour diffusion. Following data collection at beamline ID29, the crystal structures from two different yeast species were determined to 2.6 and 2.8 Å resolution, respectively. The structures not only provided the molecular details of how the kleisin subunit binds along the inner surface of the harp-shaped HEAT-repeat solenoid, but also gave a clear indication of the DNA binding groove in the complex. Mutation of conserved positively charged residues in this groove confirmed its functional importance for recruiting condensin to chromosomes both in vitro and in vivo.
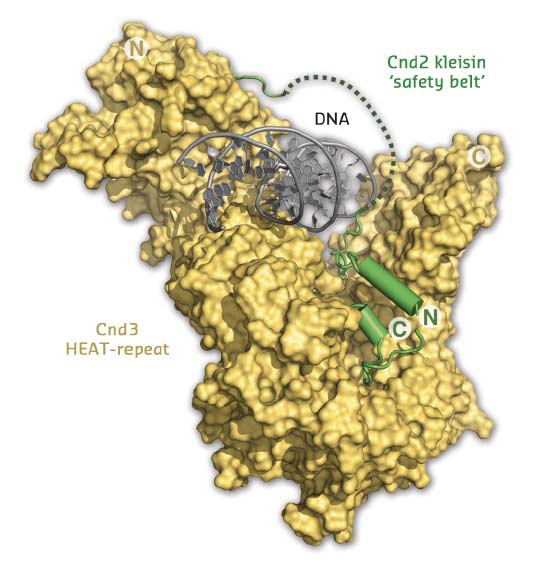
Additional extensive DNA screening yielded co-crystals of the sub-complex bound to DNA, for which data were collected at ID23-2 and a structure determined to 3.0 Å by molecular replacement (Figure 12). The resulting structure not only confirmed that the DNA double helix passes through the positively charged groove; it also suggested that the kleisin subunit encircles the bound DNA in a manner reminiscent of the way a safety belt fastens a seated passenger. Based on in vitro site-specific cross-linking and DNA binding experiments, as well as chromatin co-localisation experiments, it was possible to show that this mode of interaction provides a simple and elegant mechanism to tightly bind a DNA substrate without the need for specific interactions with the DNA bases.
 |
|
Fig. 12: Structure of the Cnd2-Cnd3 subcomplex bound to double-stranded DNA. Entrapment of the DNA helix by the Cnd2 ‘safety belt’ is indicated by a dotted line (disordered in the crystal). |
Taken together, the results provide molecular insights into the architecture of eukaryotic condensin complexes and identify a major DNA binding site, formed by Cnd3 HEAT-repeat and Cnd2 kleisin subunits in the complex. The new structures explain the preference for double-stranded DNA binding and lack of sequence specificity. The findings suggest a sophisticated DNA ‘safety belt’ mechanism that can be used to anchor condensin complexes to chromosomes, thereby enabling the formation of large chromatin loops as the fundamental organisational principle of higher-order chromatin structure.
Principal publication and authors
Structural Basis for a Safety-Belt Mechanism That Anchors Condensin to Chromosomes, M. Kschonsak (a), F. Merkel (a), S. Bisht (a), J. ;Metz (a), V. Rybin (b), M. Hassler (a) and C.H. Haering (a), Cell 171, 588-600 (2017); doi: 10.1016/j.cell.2017.09.008.
(a) Cell Biology and Biophysics Unit, Structural and Computational Biology Unit, European Molecular Biology Laboratory (EMBL) Heidelberg (Germany)
(b) Protein Expression and Purification Core Facility, European Molecular Biology Laboratory (EMBL) Heidelberg (Germany)
References
[1] W. Flemming, in Zellsubstanz, Kern und Zelltheilung (1882).
[2] Hirano et al., Cell 89, 511-521 (1997).
[3] Piazza et al., Nat. Struct. Mol. Biol. 21, 560-568 (2014).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.