- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2014
- X-ray Imaging
- Morphological evolution of the growing mollusc shell revealed by X-ray computed microtomography
Morphological evolution of the growing mollusc shell revealed by X-ray computed microtomography
Living organisms form complex mineralised biocomposites that perform a variety of essential functions. These biomaterials are often multifunctional, being responsible for not only structural support and mechanical strength, but also provide optical, magnetic or sensing capabilities. This remarkable diversity in functionality is accomplished despite a relatively narrow range of constituent inorganic materials. Hence, a significant effort has been directed to study the process of biomineralisation – to understand how organisms assimilate elements from the environment in which they live and incorporate them into living tissues. Many studies have focused on the biological and chemical aspects of the interaction between inorganic precursors and macromolecular components resulting in controlled growth of different biominerals. In contrast, thermodynamic constraints that govern many aspects of microstructure formation and morphology of the mineralised tissues are much less understood.
Mollusc shells are widely used as model systems to investigate the biomineralisation process of calcium carbonate (CaCO3) by living organisms. For example, the shell of Pinna nobilis has two juxtaposed mineralised layers with completely different properties and therefore, provides two different examples of calcium carbonate assemblies within the same organism: a calcitic layer exhibits an elongated prismatic microstructure and an aragonitic layer exhibits a ‘brick-and-mortar’ nacreous microstructure. In the prismatic layer, the mineral prism building blocks are surrounded by a 1 µm think organic interprismatic matrix defining the shape of individual prisms. The biomineralisation process of the entire prismatic layers is initiated on the organic layer covering the shell (periostracum) and propagates towards the inside of the shell by the secretion of a mixture containing the precursors for calcite formation [1]. The microstructure of the layer, a few millimetres in thickness, evolves as an assembly of growing prisms and the interprismatic matrix as a function of the growing thickness of the layer. Hence, the growth history of the prismatic layer is recorded in its microstructure and can be accessed by an appropriate imaging technique, interpreting the coordinate perpendicular to the layer as a measure of time.
The thickness of the organic interprismatic matrix surrounding the calcitic prisms matches well the resolving power accessible at ID19, the microtomography beamline. To study the shell geometry, a combination of absorption- and phase-contrast enhanced imaging modes was used to resolve the organic/calcite prism boundaries and to reconstruct the microstructure of the prismatic layer. The analysis of the 3D microtomography data was performed in the scope of classical grain growth and coarsening theories while considering the growth as a two-dimensional problem where the time parameter was replaced by the direction of the structural evolution [2]. We focused on mean field considerations, where the growth kinetics of a single prism is described by an average behaviour of the entire prismatic layer and topological considerations of space filling.
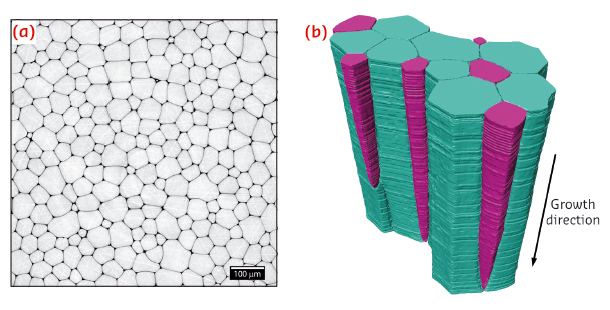 |
|
Fig. 65: a) Representative 2D microtomography section obtained perpendicular to the growth direction of the prismatic layer. b) Representative 3D microtomography section of the prismatic layer of Pinna nobilis. Growing prisms are colour-coded blue, shrinking prisms are colour-coded purple. |
Figure 65a shows a representative microtomography cross section, exhibiting a honeycomb-like microstructure in the plane perpendicular to the growth direction of the prismatic layer. A representative 3D rendering of a segment of the layer, reconstructed from a continuous set of 2D sections similar to the one presented in Figure 65a, is shown in Figure 65b. These data clearly demonstrate that the average area (cross-section) of the prisms increases during growth, in an effort to decrease the total interface area between the prisms and the adjoining organic matrix. In complete agreement with the predictions from classical grain growth theories based on mean field and on topological considerations, some of the prisms are seen to shrink and disappear while others grow in their cross-sectional area (purple and blue prisms in Figure 65b, respectively). Moreover, the data reveal that the grain size distribution renormalised by its mean value is time invariant during microstructural evolution, so that the coarsening process can be considered statistically self-similar.
This quantitative description of the structural development of the calcitic prismatic layer in P. nobilis using high-resolution microtomography and based on the comparison with well-known theories of grain growth is a fundamental step towards a better understanding of various aspects of biomineralisation. The results show how the living organism uses thermodynamic boundary conditions to define the morphology of its calcite shell.
Principal publication and authors
B. Bayerlein (a), P. Zaslansky (b), Y. Dauphin (c), A. Rack (d), P. Fratzl (a) and I. Zlotnikov (a), Nature Materials 13,1102–1107 (2014).
(a) Department of Biomaterials, Max Planck Institute of Colloids and Interfaces, Potsdam (Germany)
(b) Charité Universitätsmedizin BSRT and Julius-Wolff Institute, Berlin (Germany)
(c) Micropaléontologie, UFR TEB, Université P. & M. Curie, Paris (France)
(d) ESRF
References
[1] F. Marin, P. Narayanappa and S. Montreuil, in Molecular Biomineralization Vol. 52, W.E.G. Müller (Ed.), Springer Verlag, 353-395 (2011).
[2] H.V. Atkinson, Acta Metall. 36, 469-491 (1988).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.