- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2008
- Structural biology
- Sorting SNAREs into clathrin-coated vesicles
Sorting SNAREs into clathrin-coated vesicles
The protein and phospholipid composition of cellular membranes defines the identity of that membrane; what binds to it and what reactions occur both on its surface and inside the cavity it surrounds. Components of the cell’s limiting membrane are involved in cell signalling, homeostasis, cell contacts and recognition by the immune system. Glycosylation, proteolytic processing and degradation of transmembrane and luminal proteins occur within cells’ organelles.
Transmembrane proteins with an enormous variety of functions are moved between cell membranes in transport vesicles. Once cargo has been sorted into a forming vesicle, the vesicle buds from the donor membrane and is then transported to and fuses with the target membrane. The directionality and specificity of this transport is regulated to a large extent by members of a family of small type II transmembrane proteins called SNAREs that are found in both vesicle and target organelle membranes. After successful formation of a trans-four-helical bundle between homologous helices from a SNARE in one membrane and three SNAREs in the other, fusion of the vesicle with the target compartment can proceed. The specificity of SNARE complex formation, which leads to the specificity of vesicle/target membrane fusion, arises from the fact that only very limited combinations of SNAREs can form productive SNARE complexes. The correct distribution and localisation of SNARE proteins are therefore critical: vesicles must contain sufficient SNAREs that can engage in complex formation at the target membrane as well as SNAREs that are being recycled to their donor compartment, most likely in an inactive form.
Post-Golgi transport is mainly mediated by clathrin-coated vesicles (CCVs), whose coats are composed of an outer clathrin scaffold linked to the membrane by clathrin adaptors. Clathrin adaptors have folded domains that bind to short-lived membrane components such as phospholipid headgroups and the GTP-bound forms of small GTPase proteins of the Arf superfamily and extended, flexible regions that contain multiple short motifs, which bind to clathrin and other CCV components. At least one of a very small number of transplantable ‘tags’, a short linear sequence motif, or alternatively a covalently attached ubiquitin molecule, are found on nearly all transmembrane proteins and bind to the folded domains of the most common clathrin adaptors, so marking the transmembrane protein for incorporation as general cargo into CCVs [1]. The mechanisms by which SNAREs are selected for incorporation into CCVs were unclear, although a lot was known about how SNAREs were incorporated into ER to Golgi COPII-coated vesicles.
 |
|
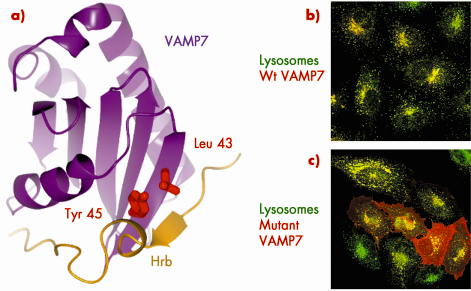
Fig. 81: a) Structure of the VAMP7 (purple): Hrb (gold) complex with key residues, whose mutation abolishes the interaction between the two proteins in vitro, highlighted in red. b) tagged VAMP7 (red) can be internalised from the cell surface to lysosomes and late endosomes (green) resulting in complete overlap of the two as indicated by the resulting yellow punctae. c) A version of VAMP7 harbouring mutations in the key VAMP7 residues Leucine43 and Tyrosine45 (red) cannot bind to Hrb and so cannot be internalised and builds up on the cell surface. |
Recent structural work carried out at beamline ID14-4 has shed light on this issue (Figure 81). The SNARE VAMP7 needs to be retrieved from the plasma membrane following its use in the fusion of late endosomes and lysosomes directly with the cell’s limiting membrane, a process thought to be important in delivering new membrane to the cell surface during membrane repair or mitosis. This retrieval occurs through the interaction of VAMP7 with the low abundance clathrin adaptor Hrb. The structure of a complex between parts of VAMP7 and Hrb showed that a 20 residue stretch of largely hydrophobic amino acids in Hrb wraps around a groove in the folded longin domain of VAMP7 that preceeds its SNARE helix. Mutation of key residues in the VAMP7/Hrb interface that were identified from the structure of the VAMP7/Hrb complex, abolish the interaction between the two proteins both in vitro and in vivo where they result in altered trafficking of VAMP7.
Taken together with other recent studies [2,3], these data on the VAMP7/Hrb interaction suggest that two parallel pathways operate in post-Golgi CCVs for cargo selection: one for selecting general cargo that is mediated by the recognition of a few widely used tags, and the other for selecting the necessary SNAREs through the recognition of determinants that are each found on the surface of only a single SNARE. By having two non-competing cargo selection systems operating simultaneously, the cell ensures that CCVs incorporate sufficient amounts of the right SNAREs to allow fusion of the vesicle with its correct target organelle as well as standard ‘bulk’ cargo.
Principal publication and authors
P.R. Pryor (a), L. Jackson (b), S.R. Gray (a), M.A. Edeling (a), A. Thompson (c), C.M. Sanderson (c,d), P.R. Evans (b), D.J. Owen (a), J.P. Luzio (a), Cell 134, 817-827 (2008).
(a) Cambridge Institute for Medical Research and Department of Clinical Biochemistry, University of Cambridge, Addenbrooke’s Hospital, Cambridge (UK)
(b) Medical Research Council Laboratory of Molecular Biology, Cambridge (UK)
(c) Medical Research Council Rosalind Franklin Centre for Genomics Research, Cambridge (UK)
(d) The Physiological Laboratory, School of Biomedical Sciences, University of Liverpool (UK)
References
[1] J.S. Bonifacino and L.M. Traub, Annu Rev Biochem 72, 395 (2003).
[2] S. Martinez-Arca, R. Rudge, M. Vacca, G. Raposo, J. Camonis, V. Proux-Gillardeaux, L. Daviet, E. Formstecher, A. Hamburger, F. Filippini, et al., PNAS 100, 9011 (2003).
[3] S.E. Miller, B.M. Collins, A.J. McCoy, M.S. Robinson, and D.J. Owen, Nature 450, 570 (2007).
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.