- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2007
- X-ray Imaging and Optics
- Phase-contrast imaging demonstrates earliest occurrence of human growth and development
Phase-contrast imaging demonstrates earliest occurrence of human growth and development
The subject of modern human origin is one of the most debated topics among anthropologists. There is little consensus about where and when the first members of our species, Homo sapiens, became fully modern. While fossil evidence tells a complex tale of mosaic change during the African Stone Age, almost nothing is known about changes in human ‘life history’, or the timing of development, reproductive scheduling, and lifespan. Tooth growth, and most importantly molar tooth eruption age, represents one of the most powerful clues to reconstruct growth processes in fossil humans [1]. Dental enamel is a unique hard tissue that begins growing early in an individual’s life, and completes formation during adolescence. It permanently records daily accretional growth (including birth), and is not subject to modification (other than wear) during an individual’s life. By using incremental growth lines in teeth, similar to annual rings in trees, developmental rate and time may be accurately established millions of years after death [2]. Recent research has shown that early fossil humans (australopithecines and early Homo) possessed short growth periods, which were more similar to chimpanzees than to living humans [3]. However, it is unclear when, and in which group of fossil humans, the modern condition of a relatively long childhood arose.
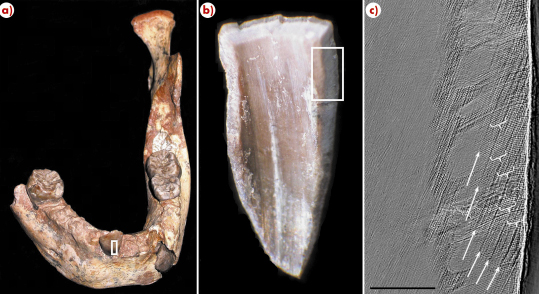
In this study, tooth growth and eruption were examined in one of the earliest representatives of Homo sapiens, found in Jebel Irhoud, Morocco and dated to approximately 160,000 years ago. With a combination of a novel application of synchrotron imaging [4-5], conventional microtomography, and developmental analysis, we reconstructed tooth formation time and determined the age at death of the fossil child (Figure 133). The results show that the Moroccan fossil, at almost 8 years of age, is currently the oldest member of the genus Homo to show a modern human life history profile. These findings and other evidence suggest that modern biological, behavioural, and cultural characteristics appeared relatively late in the past six million years of human evolution.
 |
|
Fig. 133: a) The North African juvenile fossil mandible showing the location of the incisor tooth enamel (white box) sampled at the ESRF. b) Close up of enamel fragment, with the area of interest shown in the white box. c) Synchrotron image showing growth lines (white arrows) with 10 daily lines between them (white brackets). Scale bar = 200 micrometres. |
The new application of high-resolution propagation phase contrast X-ray synchrotron local microtomography, whose first results were obtained four years ago [4], permits previously inaccessible developmental features to be revealed non-destructively [4-5]. By imaging teeth prior to physical sectioning and comparing the same areas in our second study, we demonstrated the accuracy of virtual histology (Figure 134). Ongoing applications of this technique will permit more precise assessments of mammalian evolutionary developmental biology, as well as greater understanding of the three-dimensional structure of dental tissues.
 |
|
Fig. 134: Comparison of (A) physical and (B) virtual sections of a chimpanzee molar germ, illustrating (C) correspondence between incremental features. Application of the 3D technique on an intact germ (D) and a fossil oran-utang molar (E). Scale bar = 500 micrometres in A and B, 100 micrometres in C, and 250 micrometres in D and E. |
Recent studies performed on the ESRF beamline ID19 have shown that microscopic incremental features in dental tissues can be imaged with high fidelity using phase contrast X-ray synchrotron microtomography. This technique has been applied for non-destructive and precise assessment of the age at death and length of the growth period in fossil teeth. Future non-destructive applications will reveal growth processes in additional rare human fossils, allowing finer resolution of the evolution of human life history.
References
[1] B.H. Smith, R.L. Tompkins, Annu. Rev. Anthropol. 24, 257–279 (1995).
[2] T.M. Smith, D.J. Reid, J.E. Sirianni, J. Anat. 208, 125-138 (2006).
[3] C. Dean et al. Nature 414, 628-631 (2001).
[4] P. Tafforeau, Ph.D. Thesis, Université de Montpellier II, pp. 284 (2004).
[5] P. Tafforeau et al., Applied Physics A 83, 195-202 (2006).
Principal publications and authors
T.M. Smith (a), P.T. Tafforeau (b,c), D.J. Reid (d), R. Grün (e), S. Eggins (e), M. Boutakiout (f), J.-J. Hublin (a), Proc. Natl. Acad. Sci. USA 104, 6128-6133 (2007); P. Tafforeau, T.M. Smith, J. Hum. Evol. 54 (in press).
(a) MPI for Evolutionary Anthropology, Leipzig (Germany)
(b) LGBPH, CNRS, Poitiers (France)
(c) ESRF
(d) Newcastle University (UK)
(e) RSES, Australian National University, Canberra (AZ)
(f) University Mohammed V-Agdal, Rabat (Morocco)
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.