- Home
- Users & Science
- Scientific Documentation
- ESRF Highlights
- ESRF Highlights 2004
- Structural Biology
- The Crystal Structure of Glycogen Synthase
The Crystal Structure of Glycogen Synthase
Glycogen and starch, megadalton-sized glucose polymers, are the major reservoir of readily-available energy and carbon compounds in most living organisms. In mammals, glucose uptake and utilisation are under tight control. Defects in normal glucose handling are associated with a variety of human pathologies such as glycogen storage diseases and diabetes, in which persistent hyperglycemia is correlated with an early onset and increased severity of the disease. The metabolic pathways for the catalytic conversion of glycogen involve several enzymes, among these glycogen synthase catalyses the elongation, and glycogen phosphorylase the breakdown, of a(1,4)-linked glucans. A great wealth of information has been gained on glycogen degradation through the biochemical and structural studies of glycogen and maltodextrin phosphorylases in the last 30 years [1]. These studies have shown that glycogen phosphorylase regulation is a fascinating example of covalent and non-covalent allosterism, exquisitely coordinated by hormonal control. Information describing the tertiary and quaternary structures is now available for practically all the enzymes directly involved in glycogen processing. Surprisingly, very little structural information has been reported on glycogen synthase, which catalyses glycogen elongation through the successive addition of ![]() -(1,4)-linked glucose residues to the polymer's non-reducing end, using ADP/UDP-glucose as glucose donor.
-(1,4)-linked glucose residues to the polymer's non-reducing end, using ADP/UDP-glucose as glucose donor.
We have now determined the crystal structure of the glycogen synthase from Agrobacterium tumefaciens (AtGS) at 2.3 Å resolution, both in its apo-form and in the presence of ADP. The data were collected on ID29, and the structure solved using the SAD technique (single wavelength anomalous diffraction), followed by density modification procedures. The protein is found as a dimer in the asymmetric unit, with each monomer comprised of two 'Rossmann-fold' domains (Figure 81) that are organised as in the structures of glycogen phosphorylase and other glycosyl transferases of the GT-B superfamily [2]. A deep fissure between both domains contains the catalytic centre. A major difference between mammalian (family GT3) and bacterial (GT5) glycogen synthases, is that only GT3 enzymes are tightly regulated by phosphorylation and non-covalent allosteric modulation. According to the structure-based multiple sequence alignments among GT3, GT5 and glycogen phosphorylase sequences, the regulatory phosphorylation sites in human glycogen synthase are probably located in the 25-residue N-terminal and 120-residue C-terminal insertions, outside the non-regulated GT-B core of bacterial glycogen synthases. Mutagenesis studies had previously identified two conserved Arg clusters in GT3 enzymes as part of the allosteric site(s) for both the phospho-Ser/Thr residues and the non-covalent activator Glc-6P. The equivalent residues in AtGS are located at one end of the C-terminal domain, far from the reaction centre, indicating a long-distance regulatory effect.
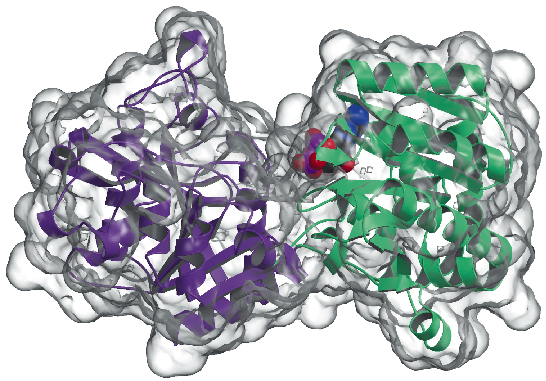 |
|
Fig. 81: Glycogen synthase structure: the solvent accessible surface is represented in transparent. Note the bound ADP molecule (drawn as VDW balls). |
The structure of AtGS shows a striking topological resemblance with the core structures of glycogen/ maltodextrin phosphorylases (family GT35) (Figure 82a), even though sequence identities are low (~14%). The structure of each Rossmann-fold domain of AtGS is significantly closer to those of phosphorylases from the GT35 family than to any other nucleotide-dependent glycosyl transferase. Glycogen synthase and phosphorylases also exhibit a strong resemblance at their catalytic sites (Figure 82b). The ADP ribose and the pyridoxal groups lie in equivalent positions, as do the distal phosphate of ADP in AtGS and the inorganic phosphate substrate in phosphorylases. Furthermore, critical amino acids that interact with the substrates' glucose moieties are also conserved. The catalytic mechanism used by glycogen synthases and phosphorylases is therefore probably similar, with subtle changes in structure explaining their overall different functional properties.
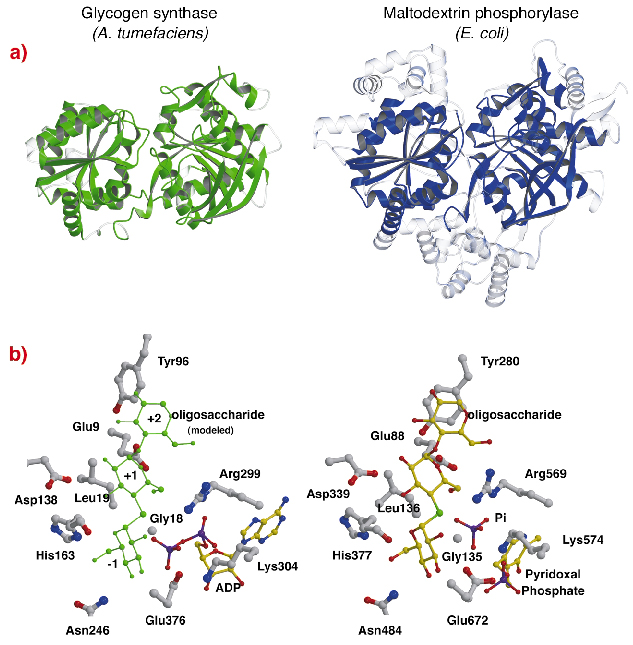 |
|
Fig. 82: Homology to glycogen phosphorylase: (a) after structural alignment, superimposed equivalent residues between AtGS and E. coli maltodextrin phosphorylase MalP, are represented in solid material (non-aligned regions, in transparent); (b) the catalytic sites of AtGS and MalP. |
The initial suggestion that glycogen synthesis simply represented a reversal of its degradative phosphorolysis was demonstrated to be flawed as Leloir showed that UDP-Glc and glycogen synthase, not Glc-1-P and glycogen phosphorylase, were responsible for polysaccharide elongation [3]. Our work now demonstrates however, that the opposite independent reactions in glucose-glycogen interconversion are catalysed by closely-related homologous enzymes.
References
[1] J.L. Buchbinder et al., Annu Rev Biophys Biomol Struct 30, 191-209 (2001).
[2] P.M. Coutinho et al., J Mol Biol 328, 307-317 (2003).
[3] L.F. Leloir and C.E. Cardini, J Am Chem Soc 79, 6340-6341 (1957).
Principal Publication and Authors
A. Buschiazzo (a), J.E. Ugalde (b), M.E. Guerin (a), W. Shepard (c), R.A. Ugalde (b) and P.M. Alzari (a) EMBO J, 23, 3196-3205 (2004).
(a) Unité de Biochimie Structurale, Inst Pasteur, Paris (France)
(b) Inst de Inv Biotecnológicas, Univ de Gral San Martín, San Martin (Argentina)
(c) ESRF
partners



European Synchrotron Radiation Facility - 71, avenue des Martyrs, CS 40220, 38043 Grenoble Cedex 9, France.